

Nature Commun | 隋森芳团队合作揭示裸藻PSI–LHCI–LHCII超复合体的独特结构与光捕获策略
光合作用是地球生物圈物质循环与能量流动的核心驱动力,为几乎所有生命活动提供能量与物质基础。藻类在全球生态系统中具有重要地位,其生物量不足高等植物的1%,却贡献了超过50%的初级生产力和氧气。真核藻类的起源被广泛认为与内共生事件密切相关。其中,绿藻门起源于蓝藻被原生动物吞噬后发生的初级内共生,其叶绿体外不包裹叶绿体内质网膜;而裸藻门、顶复门和甲藻门等类群则经历了次级内共生,其叶绿体外具有单层叶绿体内质网膜。在次级内共生过程中,常伴随类囊体膜系统的重排,这种重排对光系统及捕光体系结构的影响,长期以来缺乏直接的结构生物学证据。光系统I(Photosystem I, PSI)是执行光能转化与电子传递功能的关键分子机器。解析裸藻PSI及其捕光天线复合物的结构,有助于揭示次级内共生谱系光反应体系的组织方式与功能特征,从而深化对光合作用进化多样性的认识。
3月5日,实验室隋森芳团队与济南大学秦晓春教授与合作在Nature Communications上发表了题为The architecture and energy transfer pathways of PSI–LHCI–LHCII in the phototrophic flagellate Euglena gracilis的研究论文。该研究成功分离纯化到裸藻(Euglena gracilis)PSI–LHCI–LHCII超复合体,解析了其2.06 Å分辨率的冷冻电镜结构,揭示了这一源自次级内共生谱系的光合系统在整体架构上的独特组织模式。

结构分析显示,与高等植物和初级内共生绿藻等绿系物种相比,裸藻PSI核心复合物的亚基组成明显简化,多种在绿色谱系中高度保守的小亚基完全缺失。与此同时,其外周天线系统却显著扩展,稳定结合了多达10个捕光天线蛋白(9个LHCI和1个LHCII)。这些天线蛋白并非分布在通常认知中的PsaF/J侧,而是集中排列于PsaM一侧。这种独特的构型在目前已解析的绿系PSI–LHCI结构中尚属首例,表明裸藻PSI在进化过程中发生了体系级的重构(图1)。此外,裸藻PsaD蛋白序列中含有一段其他光合生物所不具备的特有插入片段。该结构占据了绿系物种中通常由多个小亚基共同占据的空间,并直接介导了PSI与LHCI和LHCII的相互作用。这一特化结构很可能是裸藻PSI–LHCI–LHCII超复合体发生体系级重构的重要分子基础。
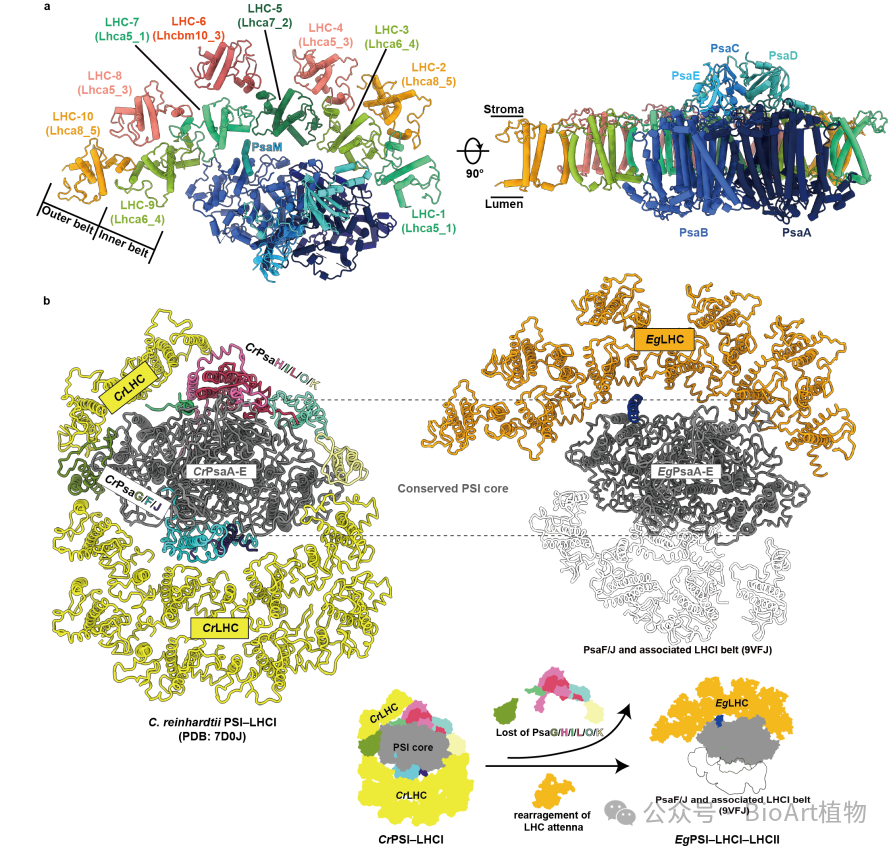
图1 裸藻PSI–LHCI–LHCII超复合体的结构组织与进化分化
在外周天线装配方面,裸藻的捕光天线系统呈现出高度有序的模块化组装特征。天线蛋白以面对面方式形成二聚体,多个LHCI–LHCI二聚体进一步组装为稳定的四聚体模块,并整体包围PSI核心。该装配方式依赖于LHCI独特的Loop_23保守结构,使得裸藻在缺失多种经典PSI小亚基的情况下,仍能维持稳定而高效的捕光体系(图2)。
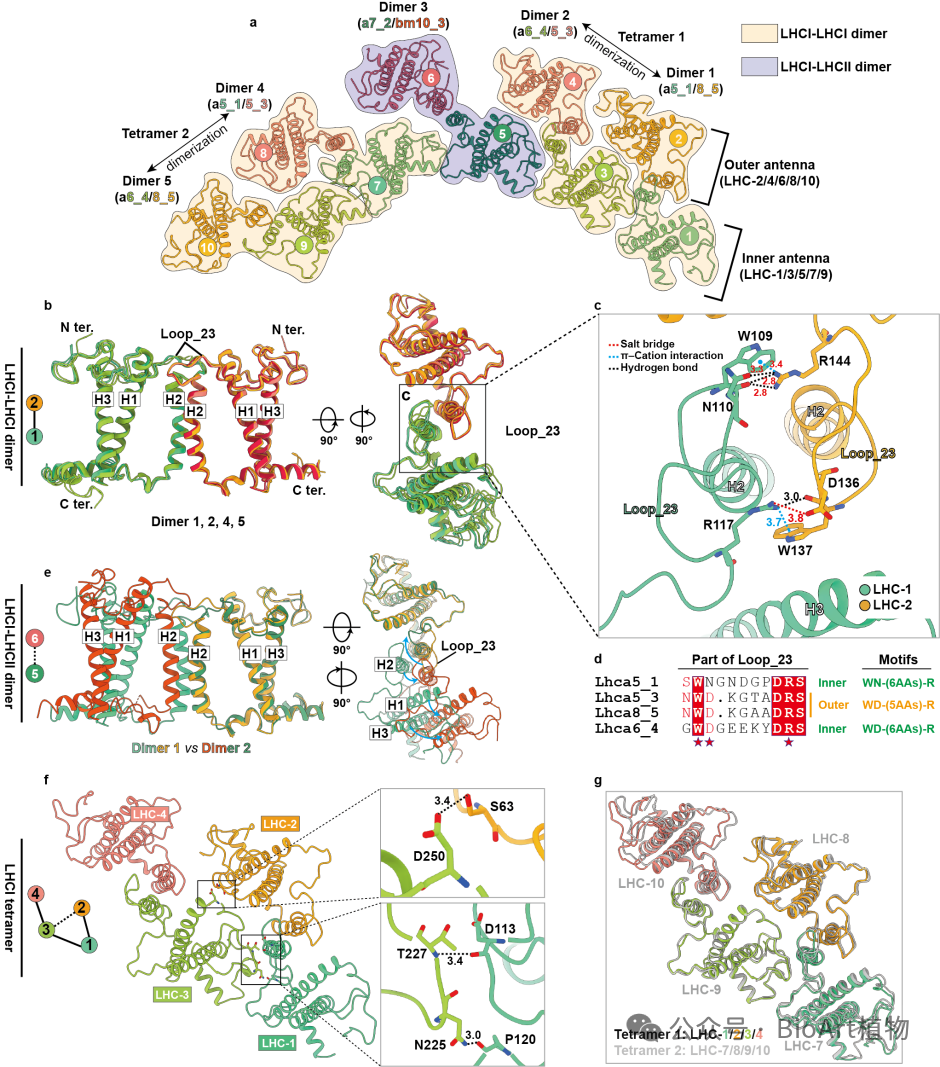
图2 裸藻PSI–LHCI–LHCII超复合体中LHC异二聚体和四聚体的层级组织
在功能层面上,裸藻PSI–LHCI–LHCII在核心与天线的连接界面处形成独特的色素网络。裸藻天线中特有的叶绿素结合位点以及红移叶绿素独特的“面对面”排列构型,共同在天线模块之间构建起高效的能量耦联通路,使激发能得以快速、定向地传递至PSI反应中心。此外,其可逆结合的LHCII体现出灵活的光适应调控策略(图3)。
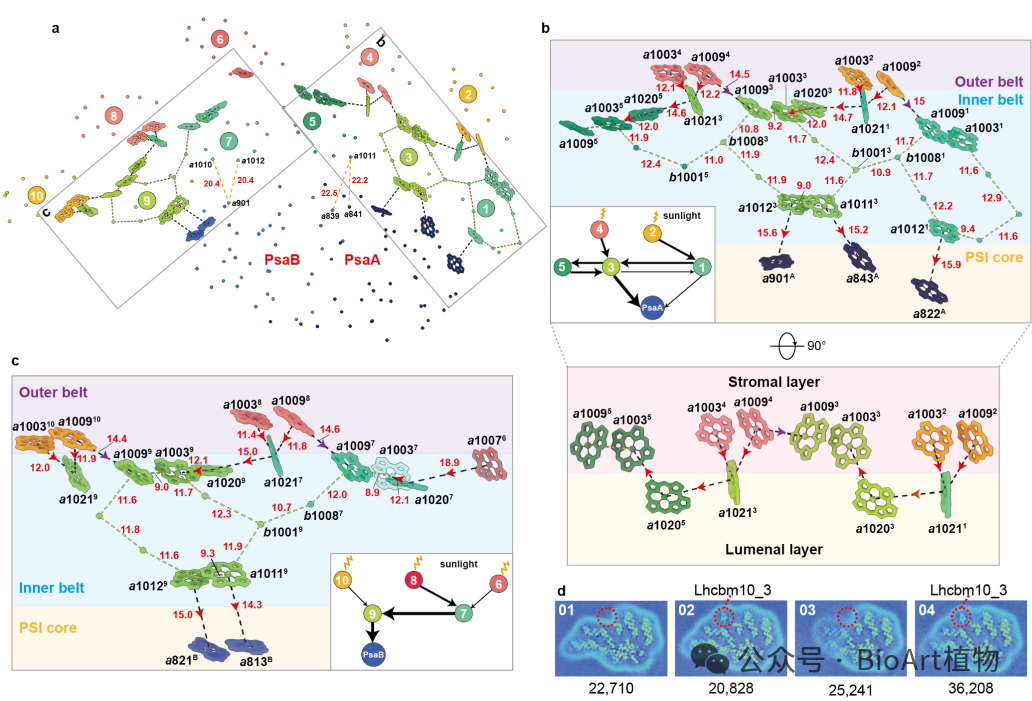
图3 裸藻PSI–LHCI–LHCII超复合体的能量传递路径
综上所述,该研究在原子分辨率水平揭示了绿系次级内共生光合生物PSI架构的独特组织方式,拓展了对光系统结构可塑性和进化多样性的理解,也为阐释天然光合系统的设计原则提供了新的结构范式。
论文链接: https://www.nature.com/articles/s41467-026-70183-y