近期研究表明TRPC5通道是治疗焦虑、抑郁和肾病的潜在药物靶点7-9。目前,由Hydra公司和Boehringer Ingelheim合作开发的一种TRPC4/5抑制剂正在临床试验中,用于治疗焦虑和抑郁10;由Goldfinch Bio开发的TRPC5抑制剂GFB-887(NCT number: NCT03970122)正处于治疗肾病的临床I期试验中。尽管TRPC5抑制剂的开发如火如荼,TRPC5抑制剂是如何结合TRPC5通道并发挥抑制作用的仍不清楚。
2021年3月8日,陈雷研究组在eLife杂志上报导了人源TRPC5通道分别与两种小分子抑制剂结合的高分辨结构。作者纯化了C端截短的人源TRPC5(hTRPC51-764),该截短体具有比全长TRPC5更高的四聚体表达量,并且像全长TRPC5一样,仍然能被激活剂(-)-englerinA(简称EA)和高浓度胞外钙激活,也能被抑制剂clemizole(简称CMZ)和HC-070抑制。CMZ是苯并咪唑衍生物,HC-070是甲基黄嘌呤衍生物,对TRPC4、TRPC5具有很高的抑制效力。这两类抑制剂具有截然不同的大小、形状和化学特性,因而可能结合在TRPC5上的不同位置。作者们向纯化后的hTRPC51-764蛋白样品中分别添加CMZ和HC-070,制备了冷冻电镜样品,通过单颗粒冷冻电镜技术获得了2.7 Å高分辨率的hTRPC51-764与CMZ结合的电子密度和hTRPC51-764与HC-070结合的电子密度并搭建了原子模型。
结构显示抑制剂结合的TRPC5具有四重对称性(图1),整个通道由跨膜区和胞质区组成,并且通道处于关闭状态。
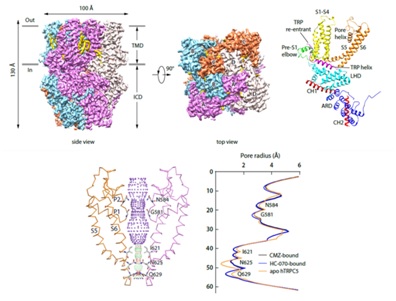
图1 抑制剂结合状态的人源TRPC5整体结构
CMZ结合在跨膜区的由跨膜螺旋S1-S4组成的电压传感器样结构域(VSLD)(图2)。与CMZ距离近的氨基酸中,F414与CMZ的氯苯环形成π-π堆积作用;Y374,M442,Y446和P659与CMZ形成疏水相互作用(图2)。功能实验结果显示,将N443突变成TRPC3/6上对应的氨基酸亮氨酸后,CMZ的抑制效力明显减弱(图2),说明N443对于CMZ发挥抑制作用很重要,也解释了CMZ对于TRPC不同亚型的选择作用。
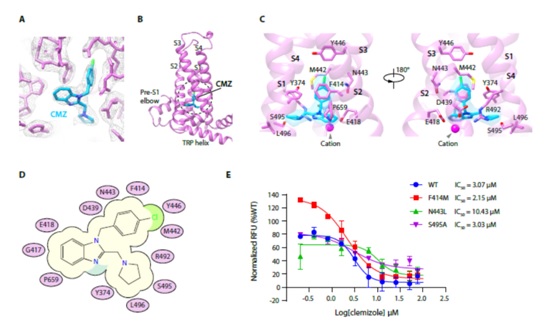
图2 CMZ在TRPC5上的结合位点
HC-070结合在跨膜区的孔道结构域,结合位点由一个亚基的跨膜螺旋S5、孔道螺旋和相邻亚基的跨膜螺旋S6组成(图3)。HC-070与TRPC5上的众多氨基酸有相互作用:F576的侧链与HC-070的甲基黄嘌呤形成π-π堆积作用;R557,Q573,W577与HC-070的羟丙基形成极性相互作用;F599, A602和T603同时也参与稳定羟丙基;F569和L572与HC-070的氯苯环形成疏水相互作用(图3)。功能实验结果显示,HC-070对以下这些突变体:R557A, F569A, L572A, Q573A, F599A,F599N,T603A的抑制效力减弱,其中,对Q573A的抑制效力减弱得最明显(图3),说明Q573A与HC-070羟丙基形成的氢键对于HC-070结合并抑制TRPC5具有至关重要的作用。
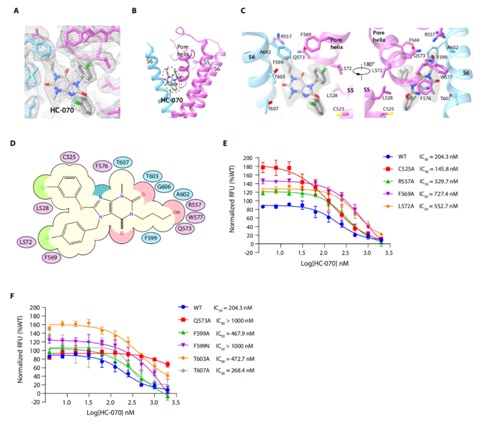
图3 HC-070在TRPC5上的结合位点
在电子密度图中,作者还观察到了结合的离子和脂类。其中一个可能的离子结合位点位于跨膜区的VSLD,由E418,E421,N436和D439组成(图4)。在鼠源TRPC4和鼠源TRPC5中也曾观察到相似密度11,12;在TRPM2,TRPM4和TRPM8中观察到的相似密度被鉴定为钙离子13-15。作者们发现E418Q-E421Q-D439N三突变体失去了对胞外钙的响应,说明该位点对于钙激活有着至关重要的作用。观察到的另一个可能的离子结合位点位于胞质区,由H172,C176,C178,C181组成,类似于细菌中葡糖激酶的HC3型锌结合位点16。C176A-C178A-C181A三突变体对EA和钙的响应不变,说明该位点可能参与离子通道门控以外的其他调控作用。
在跨膜区有数个脂类/去垢剂结合位点(图4)。在结合CMZ的TRPC5电子密度图中,在跨膜螺旋S5和S6之间观察到了类似DAG的密度。HPLC-MS实验也证明蛋白样品中含有DAG。在结合HC-070的TRPC5电子密度图中,DAG头部的密度被HC-070替代了,而两条脂肪链尾巴的密度却还保留着。
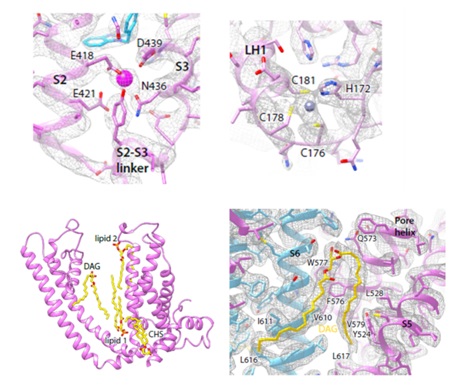
图4 离子和脂类在TRPC5上的结合位点
综上所述,本项研究解析了人源TRPC5与两类不同的抑制剂结合的高分辨结构,揭示了这两类抑制剂在TRPC5上的结合位点,并发现抑制剂通过将TRPC5稳定在无配体结合的关闭状态从而发挥其抑制功能。此项研究为深入理解TRPC5的门控机制和基于结构的药物优化奠定了基础。
本项研究的第一作者为北京大学未来技术学院分子医学所博士研究生宋康成和韦淼;北京大学分子医学研究所代谢研究检测平台主管权力博士完成了DAG的鉴定实验;博士生郭文君、康云路和博士后吴惊香参与了部分实验;陈雷研究员为通讯作者。本工作获得科技部重点研发计划、国家自然科学基金委、生命科学联合中心等经费支持。该工作的冷冻电镜样品制备、筛选和采集在北京大学电镜室和冷冻电镜平台上完成,得到了李雪梅、郭振玺、邵博、裴霞和王国鹏等人的帮助。
参考文献
1 Clapham, D. E. TRP channels as cellular sensors. Nature 426, 517-524, doi:10.1038/nature02196 (2003).
2 Philipp, S. et al. A novel capacitative calcium entry channel expressed in excitable cells. The EMBO journal 17, 4274-4282, doi:10.1093/emboj/17.15.4274 (1998).
3 Sossey-Alaoui, K. et al. Molecular Cloning and Characterization of TRPC5 (HTRP5), the Human Homologue of a Mouse Brain Receptor-Activated Capacitative Ca2+ Entry Channel. Genomics 60, 330-340, doi:https://doi.org/10.1006/geno.1999.5924 (1999).
4 Uhlén, M. et al. Tissue-based map of the human proteome. Science (New York, N.Y.) 347, 1260419, doi:10.1126/science.1260419 (2015).
5 Riccio, A. et al. Essential role for TRPC5 in amygdala function and fear-related behavior. Cell 137, 761-772, doi:10.1016/j.cell.2009.03.039 (2009).
6 Schaldecker, T. et al. Inhibition of the TRPC5 ion channel protects the kidney filter. The Journal of clinical investigation 123, 5298-5309, doi:10.1172/jci71165 (2013).
7 Just, S. et al. Treatment with HC-070, a potent inhibitor of TRPC4 and TRPC5, leads to anxiolytic and antidepressant effects in mice. PloS one 13, e0191225, doi:10.1371/journal.pone.0191225 (2018).
8 Yang, L. P. et al. Acute Treatment with a Novel TRPC4/C5 Channel Inhibitor Produces Antidepressant and Anxiolytic-Like Effects in Mice. PloS one 10, e0136255, doi:10.1371/journal.pone.0136255 (2015).
9 Zhou, Y. et al. A small-molecule inhibitor of TRPC5 ion channels suppresses progressive kidney disease in animal models. Science (New York, N.Y.) 358, 1332-1336, doi:10.1126/science.aal4178 (2017).
10 Wulff, H., Christophersen, P., Colussi, P., Chandy, K. G. &Yarov-Yarovoy, V. Antibodies and venom peptides: new modalities for ion channels. Nature Reviews Drug Discovery 18, 339-357, doi:10.1038/s41573-019-0013-8 (2019).
11 Duan, J. et al. Structure of the mouse TRPC4 ion channel. Nature communications 9, 3102, doi:10.1038/s41467-018-05247-9 (2018).
12 Duan, J. et al. Cryo-EM structure of TRPC5 at 2.8-A resolution reveals unique and conserved structural elements essential for channel function. Science advances 5, eaaw7935, doi:10.1126/sciadv.aaw7935 (2019).
13 Autzen, H. E. et al. Structure of the human TRPM4 ion channel in a lipid nanodisc. Science (New York, N.Y.) 359, 228-232, doi:10.1126/science.aar4510 (2018).
14 Huang, Y., Winkler, P. A., Sun, W., Lü, W. & Du, J. Architecture of the TRPM2 channel and its activation mechanism by ADP-ribose and calcium. Nature 562, 145-149, doi:10.1038/s41586-018-0558-4 (2018).
15 Yin, Y. et al. Structural basis of cooling agent and lipid sensing by the cold-activated TRPM8 channel. Science (New York, N.Y.) 363 (2019).
16 Miyazono, K.-i. et al. Substrate Recognition Mechanism and Substrate-Dependent Conformational Changes of an ROK Family Glucokinase from <span class="named-contentgenus-species" id="named-content-1">Streptomycesgriseus</span>. Journal of Bacteriology 194, 607, doi:10.1128/JB.06173-11 (2012).




