DNA复制起始在真核生物细胞体中受到严格精密的调控。DNA复制启动因子首先结合到DNA复制起点,以加载DNA复制解旋酶MCM2-7复合物到DNA上,随后MCM2-7被激活,DNA双链被解螺旋,从而启动DNA复制。真核生物的复制启动因子ORC由六个亚基组成。所有真核生物都是利用ORC来标记DNA复制起始的位点。除了在基因组稳定性以及肿瘤的发生发展中显而易见的重要作用,人源 ORC复合物的多个亚基(包括ORC1,ORC4,ORC6,Cdc6)的功能缺失突变都和一类罕见遗传发育疾病直接相关(Meier-Gorlin syndrome,MGS)。MGS患者在胚胎期生长迟缓并伴随多种发育畸形,出生后发育也严重受阻并导致身材矮小、小耳症、膝盖骨缺失。
虽然在不同的真核生物中,ORC的蛋白质序列高度保守,但是ORC对DNA复制起点序列的选择性在不同物种间差别很大。酿酒酵母的ORC可以识别特异的DNA复制起点,而人源细胞的ORC结合的DNA序列却没有序列特异性,主要依赖染色体结构识别复制起点。ORC序列识别差异背后的分子机制,也一直是个未解之谜。造成这种局面一个很大的原因,就是多年来一直没有ORC结合DNA的高分辨结构。已经发表的关于ORC结构中,要么复合物中没有DNA,要么分辨率较低,不足以提供足够的细节信息,我们仍然无法推测ORC与DNA是如何相互作用从而实现其功能的。为了解开这个谜团,高宁课题组和戴碧瓘课题组克服了一些技术问题,合作解析了酿酒酵母ORC结合DNA复制起始位点3-Å分辨率的冷冻电镜结构。
本文解析的ORC-ARS305 DNA复合物的结构中,ARS305包含一段ARS高度保守序列(ARS consensus sequence, ACS)和一段B1元件序列,长度为72 bp。在这个结构中,ORC的六个亚基通过与磷酸骨架的非特异性以及与碱基的特异性相互作用环绕DNA,并在ACS和B1位点使DNA发生弯曲。该结构的一个关键特征是Orc1的保守碱性氨基酸区域(Orc1-BP,basic patch)深深地插入ACS的小沟中进行序列特异的碱基识别。另外,酵母特有的具有物种特异性的位于Orc4 Wing Helix结构域(WHD)中的Helix Insertion(Orc4-IH)嵌入ACS的大沟中,与相应的碱基形成疏水相互作用。重要的是,在ACS区域形成的这些碱基特异的相互作用的碱基都非常保守。此外,在B1区域中,也有类似的来自Orc2和Orc5的碱性氨基酸区域插入到大沟和小沟中,与碱基相互作用,并使DNA弯曲。因此,酿酒酵母ORC高度序列特异性主要是通过ORC亚基的大沟、小沟插入基序与ACS保守碱基之间的特异性相互作用实现的。序列比对分析显示,所有真核生物Orc1的N端都具有类似酿酒酵母的Orc1-BP;然而Orc4-IH却只是在酿酒酵母中存在。这些发现,很大程度上解释了不同物种ORC识别起始DNA特异性差异背后的原因。
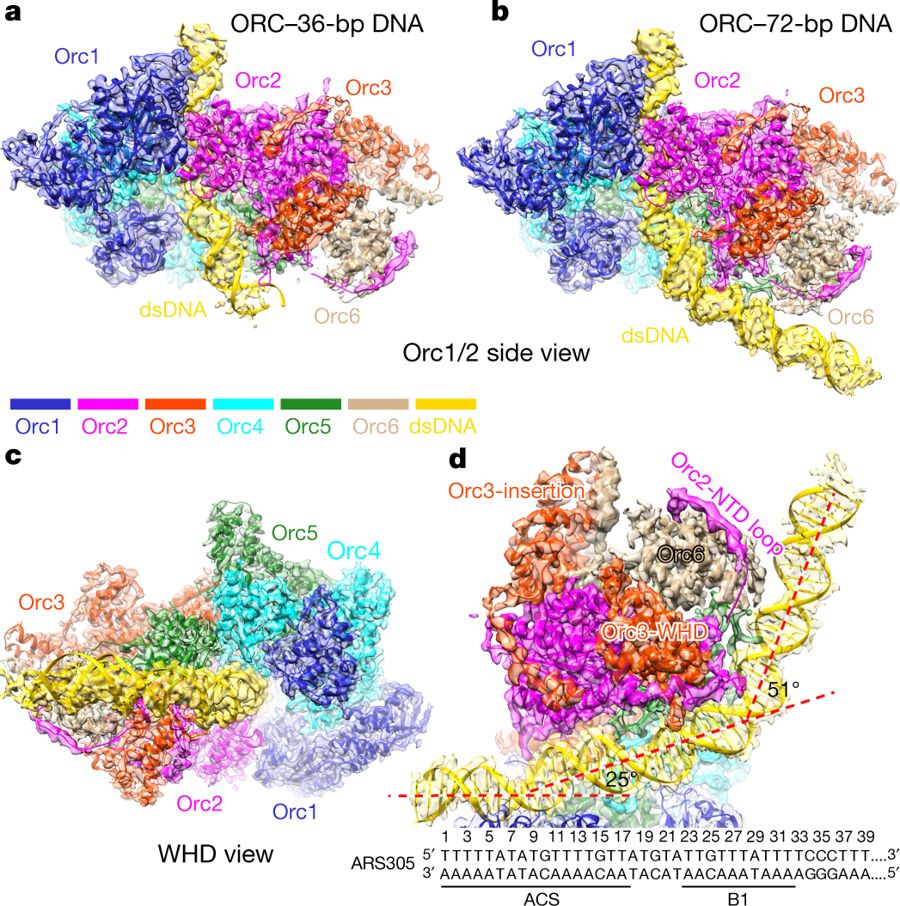
a, Side view of the cryo-EM density map of the ORC–DNA complex (36 bp) with the atomic model superimposed. dsDNA, double-stranded DNA. b, c, Side (Orc1/Orc2) (b) and bottom (WHD) (c) views of the ORC–DNA complex (72 bp). d, Cut-away view of the ORC–72-bp DNA complex, highlighting the interaction of Orc6 with Orc2 and Orc3. ARS305 sequence is shown, with successive bending of the DNA illustrated by the dashed red lines. Other Orc subunits are omitted for clarity. NTD, N-terminal domain.




